[Turku PET Centre] [PET data analysis] [Main page] [Previous page] [Next page]
All reliable information about the transport and biochemical reactions of the tracer must be incorporated in the comprehensive model. As a consequence, comprehensive model consists of many separable pools. The mathematical complexity increases drastically as the number of compartments becomes large. Analytical solutions for models of more than three compartments are possible only in rare cases; a model needs to be reduced to become a useful tracer kinetic model. However, it should be kept in mind that simplicity is no proof of validity and that validity is not necessary a prerequisite to utility [Garfinkel 1980].
There are several approaches to reduce the complexity of a model by using the quantitative information of physiology and biochemistry:
If the relative magnitudes of a series of transport steps or reactions are known, the limiting (slower) steps can be identified. Separate compartments can then be combined to form a single compartment if the transport rates between them are not limiting. For example, in brain tissue the transport across the cell membrane is assumed to be much faster than the transport across the capillary wall; concentration ratio between the interstitial space and the intracellular space is nearly in equilibrium at all times. Therefore these compartments can be approximated as a single compartment.
With ligand-receptor models, a common assumption is that the binding and release of ligand from nonspecific binding sites is not a limiting step(k5 & k6 >> k3 & k4), and the compartments of non-specifically bound and free ligand in tissue can be combined into one.
If the delivery of tracer is high compared to the extraction (PS value is small compared with blood flow), so that the plasma concentration of tracer along the capillary does not vary much, the transport of tracer into tissue is almost independent of blood flow. In such a case the model can be further simplified by eliminating the effect of blood flow. For example, the FDG model could be reduced to the configuration shown in figure 2-4.
![]()
Reduced and workable compartmental model for FDG kinetics in brain tissue.
Another approach to reduce the complexity of a model and to resolve the nonuniqueness problem is to assign parameter to a known or fixed value. Although the mathematical complexity of the model solution is not reduced, the parameter estimation problem is improved. Simpler models are more tolerant of noise and require less detailed data than more complex models, if the goal is to determine all the parameters in the more complex model. A more realistic model with some parameters pre-established at reasonable levels is more accurate than a less realistic model that ignores those parameters altogether [Graham 1985].
Fixing the distribution volume (K1/k2) for 15O labeled water for a literature value, or assigning an independently measured value for vascular fraction in tissue, are examples of this approach. In brain receptor studies, an estimate of K1/k2 in receptor-free area can sometimes be assigned for other regions.
If the assigned values of the parameters differ from the true values of a particular study, the deviation can be reflected as systematic errors in other parameters. Sometimes a trade-off has to be made: which are more useful, parameter estimates with bias or with large variability.
The physical meaning of the parameters is changed when the number of compartments in a model is reduced. When two compartments are combined to one, the rate constants for transport out of the new compartment refer to the total amount of tracer in the new compartment. This change must be considered when assigning fixed values to parameters, validating the model, or interpreting model predictions.
The value of a rate constant in a compartmental model is different from the physical of chemical rate constants. Since the rate constant in compartmental model is with respect to the amount of tracer in a compartment, it is related to both the tracer concentration and the volume of the space that the compartment represents. The term distribution volume (Vd) is frequently used in tracer kinetics; it is the volume of tissue space in which the tracer would have distributed with the same concentration as in blood. It is defined mathematically as the equilibrium ratio of the concentration of tracer in tissue to the concentration of tracer in blood, and is identical to the term partition coefficient, p.
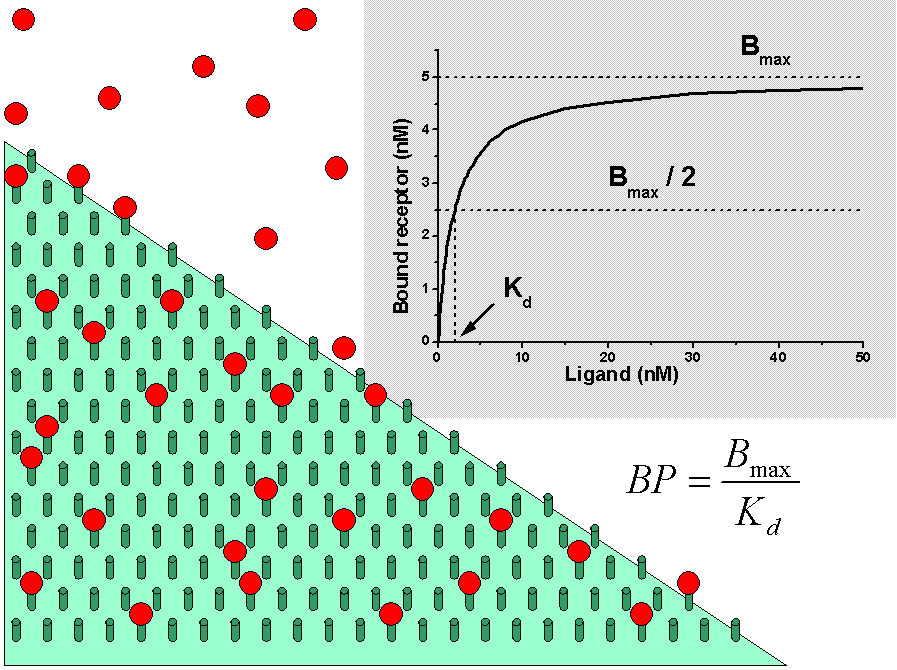
Receptor model can often be reduced to a similar model than with [18F]FDG. After combining the compartments for nonspecific binding and free ligand, the physical meaning of partition coefficient changes, and the actual value can be very different from the expected value (usually near unity). The nonspecific binding to plasma proteins is usually ignored, which also changes the partition coefficient. In receptor model with tracer concentrations the parameters k3 and k4 can be defined with terms of receptor binding:
Parameters kon and B'max (or k3 and k4) are not separable from a single bolus injection experiment under tracer conditions [Mintun et al. 1984]. Only BP (=f2B'max/KD, where f2 is the fraction of free ligand in tissue) can be determined from one kinetic study at a single condition.
Garfinkel D. Computer modeling, complex biological systems, and their simplifications. Am J Physiol 1980; 239: R1-R6.
Mintun MA, Raichle ME, Kilbourn MR, Wooten GF, Welch MJ. A quantitative model for the in vivo assessment of drug binding sites with positron emission tomography. Ann Neurol. 1984; 15: 217-227.
Phair RD. Development of kinetic models in the nonlinear world of molecular cell biology. Metabolism 1997; 46:1489-1495.